 | |
| |
 |  |  | |||||
| Präparat: 98 (L) |
| Cornea, mehrschichtiges unverhorntes Plattenepithel ohne Papillenbildung |
| H.-E. |
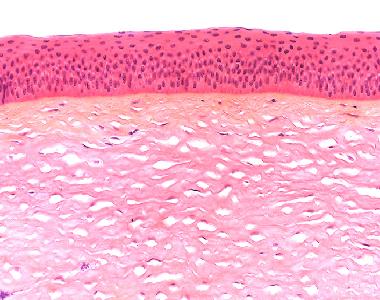 |
Das außen gelegene mehrschichtige unverhornte Plattenepithel besteht aus mehreren Zelllagen. Drei Schichten können hinsichtlich ihrer Form und Funktion unterschieden werden: Die Regeneration des Epithels erfolgt aus den hochprismatischen Zellen der basalen Schicht (Stratum basale). Daher finden sich in dieser Schicht zahlreiche Mitosen. Innerhalb von sieben Tagen wandern die neugebildeten Zellen aus der basalen in die oberflächliche Schicht. Die Zellen wandern zunächst in die Intermediärzellschicht (Stratum intermedium), wo sie polygonal geformt sind und durch Desmosomen miteinander verbunden sind. Die oberste Schicht (Stratum superficiale) besteht aus Plattenepithelzellen und läßt im Gegensatz zum mehrschichtigen verhornten Plattenepithel deutliche Zellkerne erkennen. Die abgeplatteten Zellen sind ultrastrukturell mit zahlreichen Mikrovilli besetzt, die in den Tränenfilm eintauchen. Das Hornhautepithel sitzt einer lichtmikroskopisch sichtbaren Basalmembran, der Bowman-Membran auf. Diese Membran besteht aus einer nur im Elektronenmikroskop erkennbaren Basallamina und einer 12 µm dicken spezialisierten Lamina fibroreticularis aus Kollagenfibrillen und einer dichten Interzellularsubstanz. Unterhalb des Epithels liegt das ca. 500 µm dicke corneale Stroma, die Substantia propria corneae aus kollagenem Bindegewebe mit Fibroblasten bzw. Fibrozyten und weiten Interzellullarräumen, die mit amorpher Grundsubstanz und kollagenen Fasern gefüllt sind. Die hohe Wasserbindungskapazität, die gleichmäßige Verteilung des Wasser im Stroma und die regelmäßige Ausrichtung und Dicke der Kollagenfibrillen sind Voraussetzung für die Lichtdurchlässigkeit der Cornea.
Epithel innerer Körperoberflächen, die starken mechanischen Belastungen ausgesetzt sind, z.B. Mundhöhle, Ösophagus, Analkanals und Vagina. Es besteht aus mehreren Schichten, deren Zellen unterschiedliche Formen besitzen: Die Zellen des Stratum basale sind hochprismatisch, relativ klein und dicht gepackt, die Zellen des Stratum spinosum sind polygonal geformt und besitzen stachelig erscheinende Ausläufer (daher der Begriff Stachelzellschicht. Die Basalzellschicht und die Stachelzellschicht bilden zusammen die Proliferationszellschicht. Von der Basalzellschicht aus wandern die Zellen an die Oberfläche und verändern dabei ihre Form. Die Zellen der Superfzialschicht sind abgeplattet (daher Plattenepithel). Die Zellkerne sind gut erhalten und bis in die obersten Zellschichten zu erkennen. Das Plattenepithel ist durch Bindegewebspapillen eng mit der Lamina propria verzahnt.
Basalzellschicht eines mehrschichtigen Epithelverbands. Sie grenzt an das unterliegende Bindegewebe. In ihr finden die Mitosen für den Zellersatz statt. Zusammen mit Anteilen der Intermediärzellschicht ist sie die Regenarationsschicht bzw. Proliferationszone aller mehrschichtigen Epithelien. Von hier steigen die Zellen zur Oberfläche des Epithelverbands auf und machen dabei eine Reifung durch. Die Basalzellen sitzen auf einer Basallamina, mit der sie durch Hemidesmosomen verbunden sind.
Bei mehrschichtigen Epithelien werden die einzelnen Zellschichten unterschiedlich benannt. Alle Epithelien, die mehr als 2 Zelllagen besitzen, lassen grundsätzlich eine grobe Gliederung in 3 Stockwerke erkennen: Basal-, Intermediär- und Superfizialschicht. Die Intermediärzellschicht liegt zwischen Basal- und Superfizialschicht und besteht meist aus mehreren Lagen relativ großer runder oder polygonaler Zellen. Das Stratum spinosum des mehrschichtigen Epithels wird auch als „Stachelzellschicht“ bezeichnet. Die Zellen sind über Desmosomen miteinander verbunden. Das Bild der Stachelzellen entsteht dadurch, dass durch Vergrößerung des Interzellularraumes (z.B. durch Schrumpfung des Zellleibes durch histotechnische Behandlung) die Abschnitte der mit Desmosomen besetzten Zelloberflächen deutlich hervortreten, was zu einem Eindruck von stachelförmigen Fortsätzen führt.
Im Elektronenmikroskop erkennbare Haftkomplexe zwischen zwei benachbarten (Epithel-) Zellen. Nach Form und Ausdehnung der Haftkomplexe unterscheidet man punktförmige Puncta adhaerentes, fleckförmige Maculae adhaerentes und Zonulae adhaerentes. Die letztgenannten „Gürteldesmosomen“ umgeben die Zelle bandartig. Ein Desmosom besteht aus zwei Halbdesmosomen der beiden benachbarten Zellen. Im Desmosomen-Bereich wird der 20-40 nm breite Interzellularraum von mikrofilamentärem Material durchzogen. Vom Zellinneren ziehen Intermediärfilamente des Zytoskeletts an eine Verdichtungszone (Plaque). Hier interagieren Plaque-Proteine einerseits mit den Intermediärfilamenten des Zytoskeletts und andererseits mit membrandurchspannenden Proteinen vom Typ der desmosomalen Cadherine. Desmosomen kommen vor allem in Epithelen vor, besonders zahlreich in mehrschichtigen Plattenepithelien, aber auch in nicht-epithelialen Geweben; im Herzmuskel, zwischen den follikulären dendritischen Zellen des Lymphfollikels und zwischen Arachnoidalzellen der weichen Hirnhaut.
Oberflächliche Zellage des mehrschichtigen unverhornten Plattenepithels aus abgeplatteten, kernhaltigen Zellen.
Bildet die äußere Oberfläche der Haut. Zu differenzieren sind von basal (dem Bindegewebe zugewandt) nach apikal (der Oberfläche zugewandt) folgende Schichten: Stratum basale, Stratum spinosum, Stratum granulosum, Stratum lucidum und Stratum corneum. Stratum basale und Stratum spinosum dienen der Regeneration des Epithelverbandes und werden auch als Stratum germinativum zusammengefasst. Stratum granulosum und Stratum lucidum bilden die Verhornungsschicht. Das Stratum corneum ist die verhornte Schicht, hier haben die zellen ihre Zellkerne verloren und bilden Hornplatten.
Mikrovilli sind fingerförmige Ausstülpungen der Zellmembran an der apikalen Seite von Zellen. Die einzelnen Mikrovilli haben einen Durchmesser von ca. 100 nm und sind 1-2 µm lang. Einzelne Mikrovilli sind nur elektronenmikroskopisch sichtbar. Stehen sie sehr dicht (z. B. im Epithel des darmes) sind sie auch lichtmikroskopisch als Bürstensaum an der Zelloberfläche zu erkennen. Die Ausbildung von Mikrovilli führt zu einer enormen, ca. 25-fachen Oberflächenvergrößerung der Zelle. Mikrovilli enthalten ca. 40 gebündelte, längsverlaufende Aktinfilamente, die über spezifische Proteine an der Plasmamembran befestigt sind. Nach basal sind die Aktinfilamente an dichten, horizontal verlaufenden Teilen des Zytoskeletts, dem terminalen Netz (terminal web), verankert.
Mit dem Begriff "Basalmembran" wird eine lichtmikroskopisch erkennbare Struktur zwischen einem Epithelverband und dem unterliegenden Bindegewebe bezeichnet. Sie ist nur dort zu erkennen, wo die Lamina fibroreticularis der Basallamina verstärkt ist.
Extrazellulär gelegene, in der Regel ca. 1 µm dicke Struktur, die Epithelien bzw. Endothelien mit dem angrenzenden Bindegewebe verbindet und in der Regel lichtmikroskopisch nicht dargestellt werden kann. Sie besteht in erster Linie aus Kollagen (Typ IV), Glykoproteinen und sauren Proteoglykanen. Elektronenmikroskopisch lassen sich vier Schichten unterscheiden: 1. Lamina rara externa (Lamina lucida), sie grenzt direkt an die Plasmamembran des Epithels. 2. Lamina densa, eine mäßig elektronendichte, 20-120 nm dicke Schicht mit Typ-IV-Kollagen und Laminin, Proteoglykanen und Glykoproteinen. 3. Lamina rara interna, eine nicht immer erkennbare Schicht mit Fibronektin. Lamina rara und Lamina densa werden als Basallamina im eigentlichen Sinne bezeichnet. 4. Lamina fibroreticularis, die dickste Schicht (200 -500 nm dick), verankert die Basallamina am Bindegewebe und besteht aus einem Geflecht dünner retikulärer Fasern (Kollagenfibrillen, v.a. Typ III). In einzelnen Fällen ist die Lamina fibroreticularis besonders verdickt, so dass die Grenze zwischen Epithel und Bindegewebe im Lichtmikroskop zu erkennen ist. In diesem Fall spricht man von einer Basalmembran.
Extrazellulärmatrix; bezeichnet Strukturen und Substanzen, welche die Zellzwischenräume ausfüllen. Besonders ausgeprägt ist die Extrazellulärmatrix im Binde- und Stützgewebe. Wichtige Bestandteile sind Kollagenfibrillen und elastische Fasern, Adhäsionsproteine (Fibronektine, Laminin) und die amorphe Grundsubstanz (Proteoglykane, Glykosaminoglykane)
Innere Schicht der Cornea aus Bindegewebe mit Fibroblasten, Grundsubstanz und Kollagenfasern. Nach vorne liegt ihr die Lamina limitans anterior (Bowman-Membran) auf, nach hinten die Lamina limitans posterior (Descemet-Membran). Die Cornea ist frei von Blutgefäßen.
Bindegewebe ist deutlich anders aufgebaut als Epithel. Im Epithelverband liegen die Zellen dicht an dicht, die Interzellularräume sind eng. Das Bindegewebe enthält weniger Zellen und große Interzellularräume. Die ortsständigen Zellen im Bindegewebe sind Fibroblasten/Fibrozyten. Außerdem kommen "freie" Zellen vor, die in das Bindegewebe einwandern oder es verlassen können und der Abwehr dienen. Die großen Interzellularräume des Bindegewebes enthalten Wasser und die extrazelluläre Matrix (amorphe Grundsubstanz und retikuläre, kollagene oder elastsiche Fasern), die von den ortsständigen Zellen gebildet wird. Bindegewebe ist im Körper weit verbreitet.
Ortsständige, spindelförmige Bindegewebszellen mit verzweigten Fortsätzen. Diese Zellen synthetisieren alle Substanzen der Extrazellulärmatrix des Bindegewebes. Fibroblasten haben eine hohe Syntheseleistung, ihr Zellkern ist reich an Euchromatin und erscheint in der histologischen Färbung hell. Fibrozyten sind weniger aktiv, ihr Zellkern ist Heterochromatin reich und ist deshalb kräftiger gefärbt. spindelförmige, meist plump geformte Vorstufe des Fibrozyts. Er tritt im wachsenden Bindegewebe auf und ist hochaktiv in der Bildung von Fasern und Grundsubstanz.
Ortsständige, spindelförmige Bindegewebszellen mit verzweigten Fortsätzen. Diese Zellen synthetisieren alle Substanzen der Extrazellulärmatrix des Bindegewebes. Fibroblasten haben eine hohe Syntheseleistung, ihr Zellkern ist reich an Euchromatin und erscheint in der histologischen Färbung hell. Fibrozyten sind weniger aktiv, ihr Zellkern ist Heterochromatin reich und ist deshalb kräftiger gefärbt.
Von ortsständigen Bindegewebszellen synthetisierte amorphe, farblose Substanz, welche die Interzellularrräume der Binde- und Stützgewebe ausfüllt. Sie bestimmt weitgehend die mechanischen Eigenschaften des jeweiligen Gewebes. Die Grundsubstanz enthält Proteoglykane, Glykoproteine und interstitielle Flüssigkeit. Proteoglykane sind große Moleküle deren zentraler, fadenförmiger Proteinkern mit unterschiedlich gebauten Glykosaminoglykanketten verbunden ist. Diese Glykosaminoglykane oder saure Mukopolysaccharide bestehen aus unverzweigten Disaccharidketten, repetitiven Einheiten von Aminozuckern und Uronsäuren, die stark negativ geladen sind. Beispiele: Hyaluronsäure (ohne core-Protein), Heparansulfat, Chondroitinsulfat, Dermatansulfat und Keratansulfat. Glykosaminoglykane dienen der Wasserspeicherung und sind wichtig für den Stoffaustausch, Glykoproteine beteiligen sich oft am Aufbau der Basallamina und Glykokalix (Strukturglykoproteine wie Fibronektin, Laminin).
Lassen sich mit histologischen Routinefärbungen im Lichtmikroskop darstellen. Sie setzen sich aus parallel angeordneten Kollagenfibrillen zusammen, die nur im Elektronenmikroskop erkennbar sind. Vorstufen des Kollagens (Prokollagen) werden von Fibroblasten synthetisiert und über Exozytose sezerniert. Die Bildung der Kollagenfibrillen erfolgt erst extrazellulär durch Abspaltung von Propeptiden und anschließender Quervernetzung. Kollagenfasern sind Bestandteil der Extrazellulärmatrix, ihre Anzahl und Anordnung bestimmt die Beschaffenheit des Bindegebewebes: Straffes Bindegewebe (z. B. in der Sehne) enthält zahlreiche, parallel angeordnete Kollagenfasern. Im lockeren Bindegewebe sind die Kollagenfasern weniger dicht und haben keine Vorzugsrichtung.
Nur im Elektronenmikroskop sichtbar, bestehen aus dem fibrillären Strukturprotein Kollagen, lagern sich zu Kollagenfasern zusammen, sind geformte Bestandteile der Extrazellulärmatrix. Sie kommen in Bindegewebe, Sehnen, Faszien, Bändern, Knorpel, Knochen und Zahnbein vor.