 | |
| |
 |  |  |  | ||||
| Präparat: 41 (L) |
| Thymus, adult |
| H.-E. |
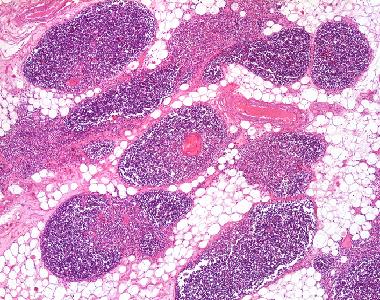 |
Eine sehr dünne und zarte Schicht von kollagenen Fasern umhüllt die einzelnen Läppchen. Die dunklere Rindenzone enthält dichtgepackte Lymphozyten mit runden, heterochromatischen Zellkernen; zwischen ihnen sind die weit weniger zahlreichen Zellkerne der epithelialen Retikulumzellen deutlich zu erkennen (größer, polygonaler, längsovaler als die Lymphozytenkerne, euchromatisch mit deutlichem Nukleolus). Das Mark der Läppchen enthält weniger dicht gepackte Lymphozyten, dazwischen ebenfalls epitheliale Retikulumzellen.
Lassen sich mit histologischen Routinefärbungen im Lichtmikroskop darstellen. Sie setzen sich aus parallel angeordneten Kollagenfibrillen zusammen, die nur im Elektronenmikroskop erkennbar sind. Vorstufen des Kollagens (Prokollagen) werden von Fibroblasten synthetisiert und über Exozytose sezerniert. Die Bildung der Kollagenfibrillen erfolgt erst extrazellulär durch Abspaltung von Propeptiden und anschließender Quervernetzung. Kollagenfasern sind Bestandteil der Extrazellulärmatrix, ihre Anzahl und Anordnung bestimmt die Beschaffenheit des Bindegebewebes: Straffes Bindegewebe (z. B. in der Sehne) enthält zahlreiche, parallel angeordnete Kollagenfasern. Im lockeren Bindegewebe sind die Kollagenfasern weniger dicht und haben keine Vorzugsrichtung.
Lappen, häufig werden Organe von aus der Kapsel einstrahlenden, mehr oder weniger derben Bindegewebszügen in Lappen und/oder Läppchen unterteilt.
Gehören zu den Leukozyten. Morphologisch können kleine und große Lymphozyten unterschieden werden. Die kleinen Lymphozyten (Durchmesser von 6-8 µm) weisen einen runden, intensiv angefärbten Zellkern auf, der von einem sehr schmalen Zytoplasmasaum umgeben ist. Funktionell und immunhistochemisch werden bei diesem Zelltyp der zellulären Immunität dienende T-Lymphozyten von antikörper-produzierenden B-Lymphozyten unterschieden. Die großen Lymphozyten haben einen Durchmesser von 8-12 µm; sie besitzen eine ovalen Zellkern. Die für T- oder B-Lymphozyten charakteristischen Oberflächenmoleküle fehlen meist, man bezeichnet sie daher auch als Non-T- bzw. Non-B-Zellen.
Kennzeichnet die aufgeknäuelten, inaktiven Chromosomenbereiche, ist in lichtmikroskopischen Präparaten intensiv angefärbt. Heterochromatin ist im Zellkern von stoffwechselinaktiven Zellen besonders ausgeprägt und liegt häufig randständig im Zellkern. Bereiche der DNA, die nicht für Proteine kodieren und stets kondensiert bleiben, wie die Telomere, werden als konstitutives Heterochromatin bezeichnet.
Dem epithelialen Anteil des Thymus zugehörige Zellen, die ein weitmaschiges Retikulum verzweigter Zellen bilden, die mit ihren langen Fortsätzen durch Desmosomen in Kontakt stehen. Charakteristisch sind die großen chromatinarmen Zellkerne.
Fasst die genetisch aktiven Abschnitte der Chromosomen zusammen, stellt die lichtmikroskopisch wenig gefärbten Regionen des Zellkerns dar. In diesen Bereichen wird die genetische Information abgelesen. Stoffwechselaktive Zellen haben einen großen, euchromatischen Zellkern.
Der Nukleolus wird auch als Kernkörperchen bezeichnet. Er stellt eine Ansammlung ribosomaler RNA und präribosomaler Partikel dar. Die Größe und die Anzahl (manchmal kommen bis zu zehn vor, i.d.R. jedoch ein bis drei) der Nukleoli ist von der Syntheseleistung der jeweiligen Zelle abhängig. In den Nukleoli findet nicht nur die Synthese der ribosomalen RNA (rRNA), sondern auch der Zusammenbau der Untereinheiten der Ribosomen statt. Auf bestimmten Nukleolus-organisierenden Chromosomenabschnitten liegen die Gene für die rRNA vor, die zur Bildung der Nukleoli führen. Hier erfolgt die Bildung einer gemeinsamen rRNA-Vorstufe, der 45-S-Prä-rRNA, aus der durch Spaltung dann die weiteren rRNA (28-S, 18-S und 5,8-S- rRNA) der Ribosomen entstehen. Elektronenmikroskopisch lassen sich innerhalb des Nukleolus drei Bereiche unterscheiden: 1. Granuläre Komponente: nimmt meist den größten Raum des Nukleolus ein; die rRNA bildet hier zusammen mit Proteinen die ribosomalen Untereinheiten (präribosomale Partikel), die als Granula vorliegen. 2. Fibrilläre Zentren: inselförmige, helle Bereiche - hier befindet sich die RNA-Polymerase I, die für die Trankription, bzw. Synthese der rRNA (45-S-Prä-rRNA) verantwortlich ist. 3. Dicht-fibrilläre Komponente: optisch dichte Bereiche, die die fibrillären Zentren schalenförmig umgeben; hier bilden spezifische Proteine Komplexe mit der Prä-rRNA, um sie anschließend in 28-S, 18-S und 5,8-S- rRNA zu zerschneiden